
Loss of Nuclear/DNA Integrity in Mouse Epididymal Spermatozoa after Short-Term Exposure to Low Doses of Dibutyl Phthalate or Bisphenol AF and Its Mitigation by Oral Antioxidant Supplementation
 , , and
, , and
Abstract
:1. Introduction
2. Materials & Methods
2.1. Animal Ethics Statement
2.2. Exposure to Pollutants
2.3. Spermatozoa Recovery
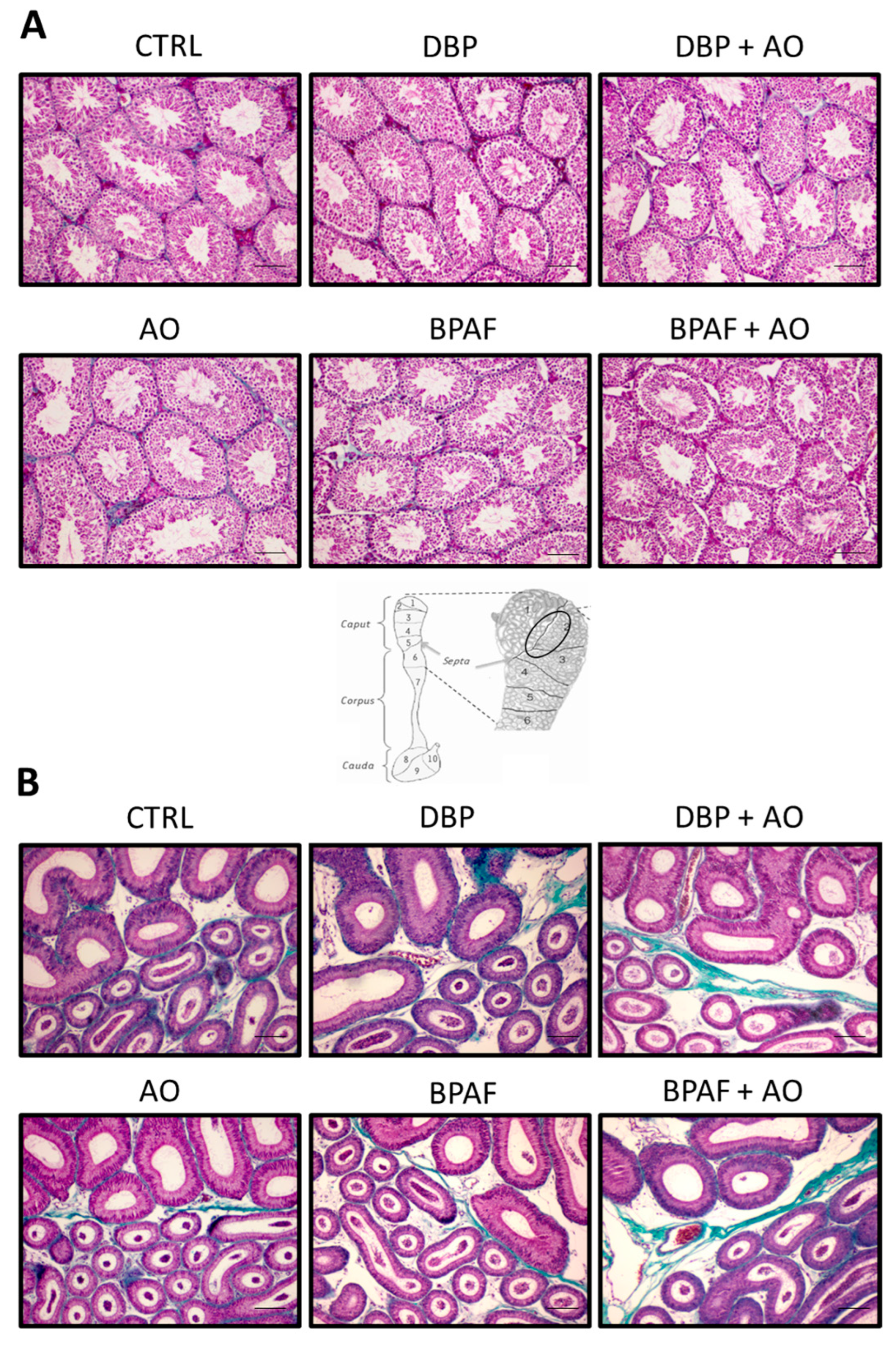
2.4. Tissue Histology
2.5. Total Protein Extraction
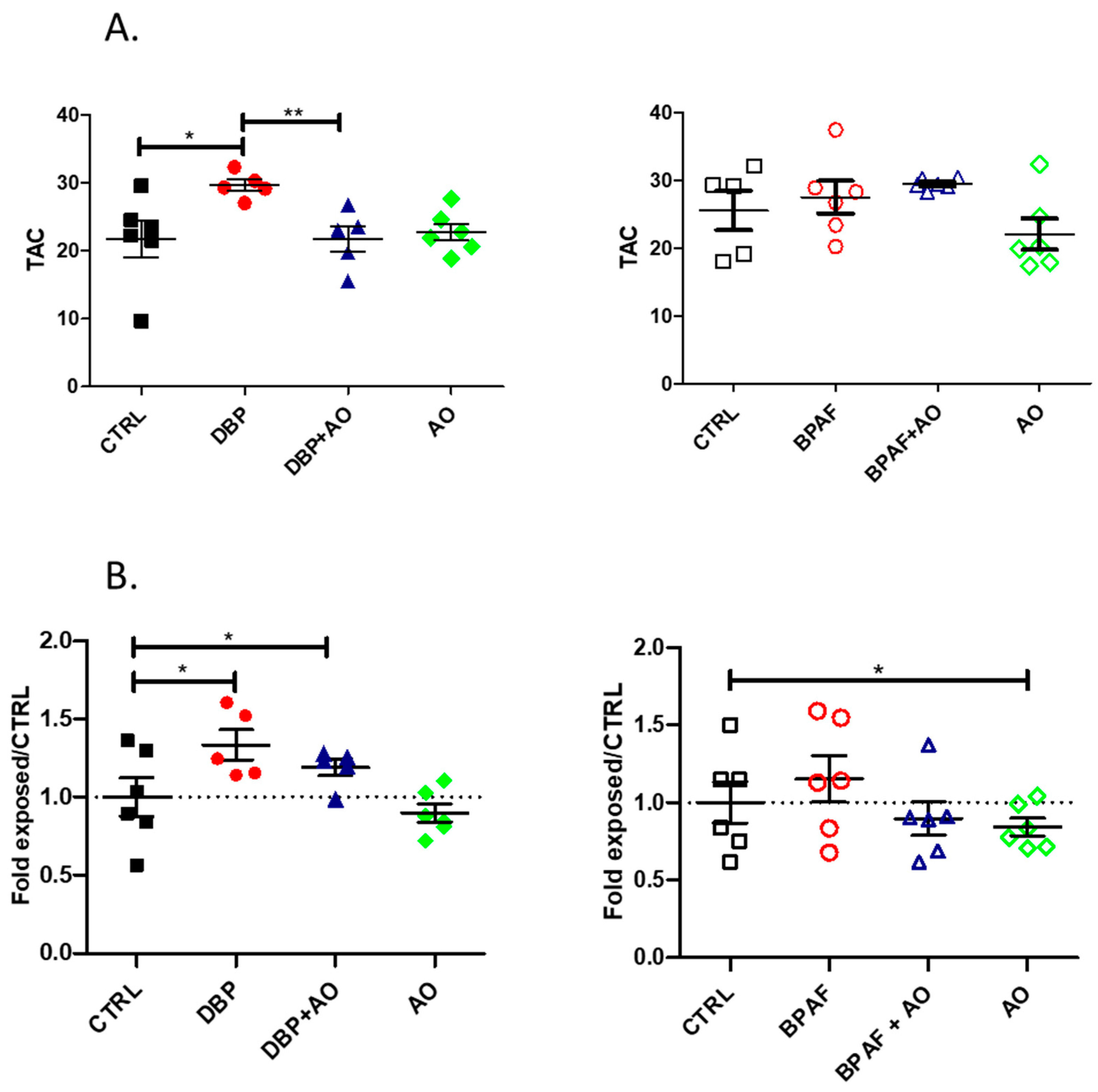
2.6. Plasma Total Antioxidant Capacity (TAC)
2.7. 4-Hydoxynonenal Analysis (4-HNE)
2.8. Standard Sperm Parameters (Mobility, Viability and Acrosome Integrity) Assessment
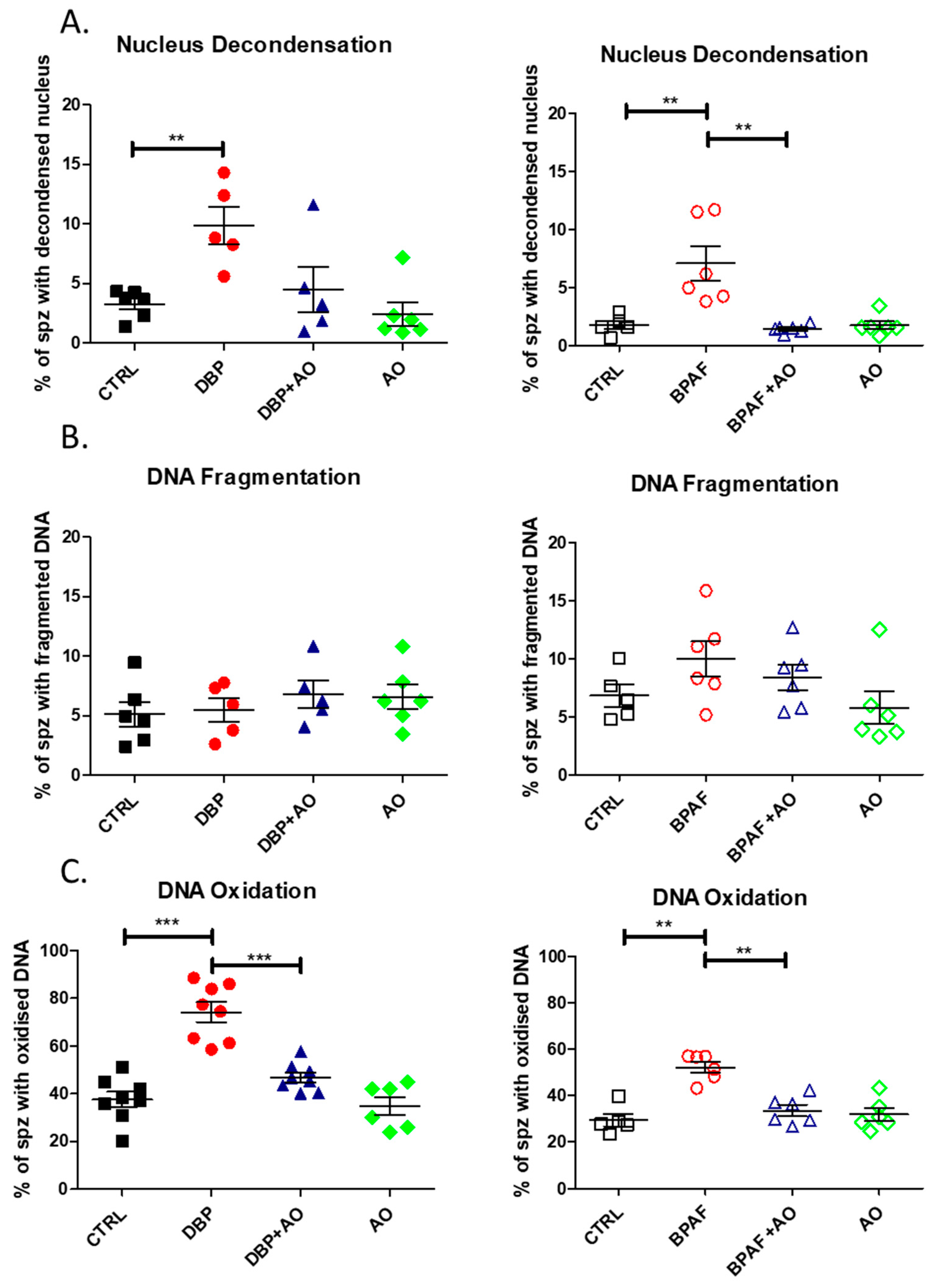
2.9. Sperm Nuclear and DNA Integrity Tests
2.10. Statistics
3. Results
3.1. Short-Term, Low-Dose Exposure to DBP or BPAF Did Not Affect the Histological Characteristics of the Testis or Epididymis
3.2. Short-Term, Low-Dose Exposure to DBP or BPAF Triggered Oxidative Stress
3.3. Short-Term, Low-Dose Exposure to DBP or BPAF Did Not Affect Classical Sperm Parameters
3.4. Short-Term, Low-Dose Exposure to DBP or BPAF Significantly Altered Sperm DNA Integrity
4. Discussion
5. Conclusions
6. Study Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Levine, H.; Jørgensen, N.; Martino-Andrade, A.; Mendolia, J.; Weksler-Derri, D.; Jolles, M.; Pinotti, R.; Swan, S.H. Temporal trends in sperm count: A systematic review and meta-regression analysis of samples collected globally in the 20th and 21st centuries. Hum. Reprod. Update 2023, 29, 157–176. [Google Scholar] [CrossRef] [PubMed]
- Levine, H.; Jørgensen, N.; Martino-Andrade, A.; Mendolia, J.; Weksler-Derri, D.; Mindilis, I.; Pinotti, R.; Swan, S.H. Temporal trends in sperm count: A systematic review and meta-regression analysis. Hum. Reprod. Update 2017, 23, 646–659. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, N.; Lamb, D.J.; Levine, H.; Pastuszak, A.W.; Sigalos, J.T.; Swan, S.H.; Eisenberg, A.W. Are worldwide sperm counts declining? Fertil. Steril. 2021, 116, 1457–1463. [Google Scholar] [CrossRef]
- Carlsen, E.; Giwercman, A.; Keiding, N.; Skakkebaek, N.E. Evidence for decreasing quality of semen during past 50 years. BMJ 1992, 305, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, P.; Dutta, S.; Krajewska-Kulak, E. The Disappearing Sperms: Analysis of Reports Published Between 1980 and 2015. Am. J. Mens Health 2017, 11, 1279–1304. [Google Scholar] [CrossRef]
- Swan, S.H.; Elkin, E.P. Declining semen quality: Can the past inform the present? BioEssays 1999, 21, 614–621. [Google Scholar] [CrossRef]
- Aitken, R.J. Role of sperm DNA damage in creating de-novo mutations in human offspring: The “post-meiotic oocyte collusion” hypothesis. Reprod. Biomed. Online 2022, 45, 109–124. [Google Scholar] [CrossRef]
- Skakkebaek, N.E.; Rajpert-De Meyts, E.; Buck Louis, G.M.; Toppari, J.; Andersson, A.M.; Eisenberg, M.L.; Jensen, T.K.; Jørgensen, N.; Swan, S.H.; Sapra, K.J.; et al. Male reproductive disorders and fertility trends: Influences of environment and genetic susceptibility. Physiol. Rev. 2016, 96, 55–97. [Google Scholar] [CrossRef]
- Skakkebaek, N.E.; Lindhal-Jacobsen, R.; Levine, H.; Andersson, A.-M.; Jørgensen, N.; Main, K.M.; Lidegaard, O.; Priskorn, L.; Holmboe, S.A.; Bräuner, E.V.; et al. Environmental factors in declining human fertility. Nat. Rev. Endocrinol. 2022, 18, 139–157. [Google Scholar] [CrossRef]
- Cannarella, R.; Gül, M.; Rambhatla, A.; Agarwal, A. Temporal decline of sperm concentration: Role of endocrine disruptors. Endocrine 2023, 79, 1–16. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Fourth Report on Human Exposure to Environmental Chemicals, Updated Tables. Atlanta, GA: U.S. Department of Health and Human Services, Centers for Disease Control and Prevention. 2018. Available online: https://www.cdc.gov/exposurereport/ (accessed on 4 September 2018).
- Teuten, E.L.; Saquing, J.M.; Knappe, D.R.U.; Barlaz, M.A.; Jonsson, S.; Bjorn, A.; Rowland, S.J.; Thompson, R.C.; Galloway, T.S.; Yamashita, R.; et al. Transport and release of chemicals from plastics to the environment and wildlife. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 2027–2045. [Google Scholar] [CrossRef] [PubMed]
- Warner, G.R.; Flaws, J.A. Bisphenol A and Phthalates: How environmental chemicals are reshaping toxicology. Toxicol. Sci. 2018, 166, 246–249. [Google Scholar] [CrossRef] [PubMed]
- Fujii, M.; Shinohara, N.; Lim, A.; Otake, T.; Kumagai, K.; Yanagisawa, Y. A study on emission of phthalate esters from plastic materials using a passive flux sampler. Atmos. Environ. 2003, 37, 5495–5504. [Google Scholar] [CrossRef]
- Lee, B.M.; Koo, H.J. Hershberger Assay for Antiandrogenic Effects of Phthalates. J. Toxicol. Environ. Health A 2007, 70, 1365–1370. [Google Scholar] [CrossRef]
- Phthalates—ECHA. Available online: https://echa.europa.eu/fr/hot-topics/phthalates (accessed on 27 March 2023).
- Aly, H.A.; Hassan, M.H.; El-Beshbishy, H.A.; Alahdal, A.M.; Osman, A.M.M. Dibutyl phthalate induces oxidative stress and impairs spermatogenesis in adult rats. Toxicol. Ind. Health 2016, 32, 1467–1477. [Google Scholar] [CrossRef]
- Awny, M.M.; Al-Mokaddem, A.K.; Ali, B.M. Mangiferin mitigates di-(2-ethylhexyl) phthalate-induced testicular injury in rats by modulating oxidative stress-mediated signals, inflammatory cascades, apoptotic pathways, and steroidogenesis. Arch. Biochem. Biophys. 2021, 711, 108982. [Google Scholar] [CrossRef]
- Yi, W.E.I.; Xiang-Liang, T.; Yu, Z.; Bin, L.; Lian-Ju, S.; Chun-Lan, L.; Tao, L.I.N.; Da-Wei, H.E.; Shang-De, W.U.; Guang-Hui, W.E.I. DEHP exposure destroys blood-testis barrier (BTB) integrity of immature testes through excessive ROS-mediated autophagy. Genes Dis. 2018, 5, 263–274. [Google Scholar] [CrossRef]
- Lyons, G. BISPHENOL A—A known endocrine disruptor. W.W.F. Eur. Toxics Programme Rep. 2000, 37. [Google Scholar]
- Goodson, A.; Summerfield, W.; Cooper, I. Survey of bisphenol A and bisphenol F in canned foods. Food Addit. Contam. 2002, 19, 796–802. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Hauser, R.; Marcus, M.; Olea, N.; Welshons, W.V. Human exposure to bisphenol A (BPA). Reprod. Toxicol. 2007, 24, 139–177. [Google Scholar] [CrossRef]
- Rochester, J.R. Bisphenol A and human health: A review of the literature. Reprod. Toxicol. 2013, 42, 132–155. [Google Scholar] [CrossRef]
- Li, D.K.; Zhou, Z.; Miao, M.; He, Y.; Wang, J.; Ferber, J.; Herrinton, L.J.; Gao, E.; Yuan, W. Urine bisphenol-A (BPA) level in relation to semen quality. Fertil. Steril. 2011, 95, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Tohei, A.; Suda, S.; Taya, K.; Hashimoto, T.; Kogo, H. Bisphenol A inhibits testicular functions and increases luteinizing hormone secretion in adult male rats. Exp. Biol. Med. 2001, 226, 216–221. [Google Scholar] [CrossRef]
- Liu, C.; Duan, W.; Li, R.; Xu, S.; Zhang, L.; Chen, C.; He, M.; Lu, Y.; Wu, H.; Pi, H.; et al. Exposure to bisphenol A disrupts meiotic progression during spermatogenesis in adult rats through estrogen-like activity. Cell Death Dis. 2013, 4, e676. [Google Scholar] [CrossRef] [PubMed]
- Babu, S.; Uppu, S.; Claville, M.O.; Uppu, R.M. Prooxidant actions of bisphenol A (BPA) phenoxyl radicals: Implications to BPA-related oxidative stress and toxicity. Toxicol. Mech. Methods 2013, 23, 273–280. [Google Scholar] [CrossRef]
- Chitra, K.C.; Latchoumycandane, C.; Mathur, P.P. Induction of oxidative stress by bisphenol A in the epididymal sperm of rats. Toxicology 2003, 185, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Bisphenol AF FDS. Available online: https://www.sigmaaldrich.com/FR/fr/sds/sial/90477 (accessed on 27 March 2023).
- Wu, D.; Huang, C.J.; Jiao, X.F.; Ding, Z.M.; Zhang, S.X.; Miao, Y.L.; Huo, L.J. Bisphenol AF compromises blood-testis barrier integrity and sperm quality in mice. Chemosphere 2019, 237, 124410. [Google Scholar] [CrossRef]
- Brassea-Pérez, E.; Hernández-Camacho, C.J.; Labrada-Martagón, V.; Vázquez-Medina, J.P.; Gaxiola-Robles, R.; Zenteno-Savín, T. Oxidative stress induced by phthalates in mammals: State of the art and potential biomarkers. Environ. Res. 2022, 206, 112636. [Google Scholar] [CrossRef]
- Sakaue, M.; Ohsako, S.; Ishimura, R.; Kurosawa, S.; Kurohmaru, M.; Hayashi, Y.; Aoki, Y.; Yonemoto, J.; Tohyama, C. Bisphenol-A affects spermatogenesis in the adult rat even at a low dose. J. Occup. Health 2001, 43, 185–190. [Google Scholar] [CrossRef]
- Dobrzyńska, M.M. Phthalates—Widespread occurrence and the effect on male gametes. Part 2. The effects of phthalates on male gametes and on the offspring. Rocz. Panstw. Zakl. Hig. 2016, 67, 209–221. [Google Scholar]
- Meeker, J.D.; Ehrlich, S.; Toth, T.L.; Wright, D.L.; Calafat, A.M.; Trisini, A.T.; Ye, X.; Hauser, R. Semen quality and sperm DNA damage in relation to urinary bisphenol A among men from an infertility clinic. Reprod. Toxicol. 2010, 30, 532–539. [Google Scholar] [CrossRef]
- Damon-Soubeyrand, C.; Bongiovanni, A.; Chorfa, A.; Goubely, C.; Pirot, N.; Pardanaud, L.; Pibouin-Fragner, L.; Vachias, C.; Bravard, S.; Guiton, R.; et al. Three-dimensional imaging of vascular development in the mouse epididymis: A prerequisite to better understand the post-testicular immune context of spermatozoa. BioRxiv 2022. [Google Scholar] [CrossRef]
- Drevet, J.R.; Hallak, J.; Nasr-Esfahani, M.H.; Aitken, R.J. Reactive Oxygen Species and Their Consequences on the Structure and Function of Mammalian Spermatozoa. Antioxid. Redox Signal 2022, 37, 481–500. [Google Scholar] [CrossRef]
- Aitken, R.J.; Drevet, J.R.; Moazamian, A.; Gharagozloo, P. Male infertility and oxidative stress: A focus on the underlying mechanisms. Antioxidants 2022, 11, 306. [Google Scholar] [CrossRef]
- Gharagozloo, P.; Gutiérrez-Adán, A.; Champroux, A.; Noblanc, A.; Kocer, A.; Calle, A.; Perez-Cerezales, S.; Pericuesta, E.; Polhemus, A.; Moazamian, A.; et al. A novel antioxidant formulation designed to treat male infertility associated with oxidative stress: Promising preclinical evidence from animal models. Hum. Reprod. 2016, 31, 252–262. [Google Scholar] [CrossRef]
- Diniz, A.; Alves, M.G.; Candeias, E.; Duarte, A.I.; Moreira, P.I.; Silva, B.M.; Oliveira, P.F.; Rato, L. Type 2 Diabetes induces a pro-oxidative environment in rat epididymis by disrupting SIRT1/PGC-1α/SIRT3 pathway. Int. J. Mol. Sci. 2022, 23, 8912. [Google Scholar] [CrossRef] [PubMed]
- Conrad, M.; Moreno, S.G.; Sinowatz, F.; Ursini, F.; Kölle, S.; Roveri, A.; Brielmeier, M.; Wurst, W.; Maiorino, M.; Bornkamm, G.W. The nuclear form of phospholipid hydroperoxide glutathione peroxidase is a protein thiol peroxidase contributing to sperm chromatin stability. Mol. Cell Biol. 2005, 25, 7637–7644. [Google Scholar] [CrossRef]
- Li, M.W.; Lloyd, K.C.K. DNA fragmentation index (DFI) as a measure of sperm quality and fertility in mice. Sci. Rep. 2020, 10, 3833. [Google Scholar] [CrossRef]
- Vorilhon, S.; Brugnon, F.; Kocer, A.; Dollet, S.; Bourgne, C.; Berger, M.; Janny, L.; Pereira, B.; Aitken, R.J.; Moazamian, A.; et al. Accuracy of human sperm DNA oxidation quantification and threshold determination using an 8-OHdG immuno-detection assay. Hum. Reprod. 2018, 33, 553–562. [Google Scholar] [CrossRef]
- Luo, G.; Wei, R.; Wang, S.; Wang, J. Paternal bisphenol a diet changes prefrontal cortex proteome and provokes behavioral dysfunction in male offspring. Chemosphere 2017, 184, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, J.; Qu, Z.; Qian, H.; Zhang, J.; Wang, H.; Xu, X.; Liu, S. Intrauterine exposure to low-dose DBP in the mice induces obesity in offspring via suppression of UCP1 mediated ER stress. Sci. Rep. 2020, 10, 16360. [Google Scholar] [CrossRef]
- Zhou, D.; Wang, H.; Zhang, J.; Gao, X.; Zhao, W.; Zheng, Y. Di-n-Butyl phthalate (DBP) exposure induces oxidative damage in testes of adult rats. Syst. Biol. Reprod. Med. 2010, 56, 413–419. [Google Scholar] [CrossRef]
- Mitsuhashi, M.; Morimura, K.; Wanibuchi, H.; Wanibuchi, H.; Hayashi, S.; Kiyota, A.; Wada, S.; Nakatani, T.; Fukushima, S. Di-n-butyl phthalate is toxic to the male reproductive system and its toxicity is enhanced by thioacetamide induced liver injury. J. Toxicol. Pathol. 2004, 17, 177–185. [Google Scholar] [CrossRef]
- Zarkovic, N. 4-Hydroxynonenal as a bioactive marker of pathophysiological processes. Mol. Aspects Med. 2003, 24, 281–291. [Google Scholar] [CrossRef]
- Prados, J.; Stenz, L.; Somm, E.; Stouder, C.; Dayer, A.; Paoloni-Giacobino, A. Prenatal exposure to DEHP affects spermatogenesis and sperm DNA methylation in a strain-dependent manner. PLoS ONE 2015, 10, e0132136. [Google Scholar] [CrossRef]
- Mokra, K.; Woźniak, K.; Bukowska, B.; Sicińska, P.; Michałowicz, J. Low-concentration exposure to BPA, BPF and BPAF induces oxidative DNA bases lesions in human peripheral blood mononuclear cells. Chemosphere 2018, 201, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.; Amaral, C.; Correia-da-Silva, G.; Almada, M.; Borges, M.; Cunha, S.C.; Fernandes, J.O.; Teixeira, N. Bisphenols A, F, S and AF trigger apoptosis and/or endoplasmic reticulum stress in human endometrial stromal cells. Toxicology 2022, 478, 153282. [Google Scholar] [CrossRef]
- Liang, S.; Yin, L.; Shengyang-Yu, K.; Hofmann, M.C.; Yu, X. High-content analysis provides mechanistic insights into the testicular toxicity of bisphenol A and selected analogues in mouse spermatogonial cells. Toxicol. Sci. 2017, 155, 43–60. [Google Scholar] [CrossRef] [PubMed]
- Harnett, K.G.; Chin, A.; Schuh, S.M. BPA and BPA alternatives BPS, BPAF, and TMBPF, induce cytotoxicity and apoptosis in rat and human stem cells. Ecotoxicol. Environ. Saf. 2021, 216, 112210. [Google Scholar] [CrossRef]
- Aitken, R.J.; Drevet, J.R. The importance of oxidative stress in determining the functionality of mammalian spermatozoa: A two-edged sword. Antioxidants 2020, 9, 111. [Google Scholar] [CrossRef]
- Drevet, J.R.; Aitken, R.J. Oxidation of sperm nucleus in mammals: A physiological necessity to some extent with adverse impacts on oocyte and offspring. Antioxidants 2020, 9, 95. [Google Scholar] [CrossRef] [PubMed]
- Chabory, E.; Damon, C.; Lenoir, A.; Kaulselmann, G.; Kern, H.; Zevnik, B.; Garrel, C.; Saez, F.; Cadet, R.; Henry-Berger, J.; et al. Epididymis seleno-independent glutathione peroxidase 5 maintains sperm DNA integrity in mice. J. Clin. Investig. 2009, 22. [Google Scholar] [CrossRef] [PubMed]
- Wood, M.L.; Esteve, A.; Morningstar, M.L.; Kuziemko, G.M.; Essigmann, J.M. Genetic effects of oxidative DNA damage: Comparative mutagenesis of 7,8-dihydro-8-oxoguanine and 7,8-dihydro-8-oxoadenine in Escherichia coli. Nucleic Acids Res. 1992, 20, 6023–6032. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Exposure | SC (M/mL) | SV | SM | SPM | SA |
|---|---|---|---|---|---|
| Control | 16.0 ± 2.9 | 38.6 ± 2.3 | 21.2 ± 4.1 | 15.6 ± 3.9 | 42.3 ± 1.6 |
| DBP | 18.6 ± 0.9 | 38.7 ± 2.4 | 22.3 ± 2.5 | 16.8 ± 2.4 | 51.1 ± 4.4 |
| DBP + AO | 18.4 ± 2.0 | 38.6 ± 2.0 | 30.5 ± 3.6 | 26.5 ± 3.8 (a) | 57.1 ± 5.2 |
| AO | 16.9 ± 1.8 | 41.7 ± 1.9 | 22.8 ± 1.8 | 16.7 ± 1.6 | 49.4 ± 2.2 |
| Exposure | SC (M/mL) | SV | SM | SPM | SA |
|---|---|---|---|---|---|
| Control | 19.4 ± 2.2 | 48.6 ± 2.4 | 31.6 ± 2.8 | 25.2 ± 1.5 | 49.6 ± 8.1 |
| BPAF | 11.7 ± 1.4 (a) | 46.8 ± 1.0 | 26.4 ± 2.6 | 21.2 ± 2.4 | 46.0 ± 4.7 |
| BPAF + AO | 12.0 ± 1.1 (a) | 46.5 ± 2.6 | 28.2 ± 4.4 | 25.3 ± 4.5 | 40.7 ± 3.0 |
| AO | 19.7 ± 1.5 | 55.0 ± 2.7 | 34.6 ± 4.2 | 23.0 ± 5.3 | 59.6 ± 5.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hug, E.; Villeneuve, P.; Bravard, S.; Chorfa, A.; Damon-Soubeyrand, C.; Somkuti, S.G.; Moazamian, A.; Aitken, R.J.; Gharagozloo, P.; Drevet, J.R.; et al. Loss of Nuclear/DNA Integrity in Mouse Epididymal Spermatozoa after Short-Term Exposure to Low Doses of Dibutyl Phthalate or Bisphenol AF and Its Mitigation by Oral Antioxidant Supplementation. Antioxidants 2023, 12, 1046. https://doi.org/10.3390/antiox12051046
Hug E, Villeneuve P, Bravard S, Chorfa A, Damon-Soubeyrand C, Somkuti SG, Moazamian A, Aitken RJ, Gharagozloo P, Drevet JR, et al. Loss of Nuclear/DNA Integrity in Mouse Epididymal Spermatozoa after Short-Term Exposure to Low Doses of Dibutyl Phthalate or Bisphenol AF and Its Mitigation by Oral Antioxidant Supplementation. Antioxidants. 2023; 12(5):1046. https://doi.org/10.3390/antiox12051046
Chicago/Turabian StyleHug, Elisa, Pauline Villeneuve, Stephanie Bravard, Areski Chorfa, Christelle Damon-Soubeyrand, Stephen G. Somkuti, Aron Moazamian, R. John Aitken, Parviz Gharagozloo, Joël R. Drevet, and et al. 2023. "Loss of Nuclear/DNA Integrity in Mouse Epididymal Spermatozoa after Short-Term Exposure to Low Doses of Dibutyl Phthalate or Bisphenol AF and Its Mitigation by Oral Antioxidant Supplementation" Antioxidants 12, no. 5: 1046. https://doi.org/10.3390/antiox12051046